Division-Filicophyta, Class-Leptosporangiopsida ve Order-Filicales (7002 Kelime)
Division-Filicophyta, Class-Leptosporangiopsida ve Order-Filicales ile ilgili faydalı notlar!
Karakteristik özellikler:
Onlar homosporlu eğrelti otlarıdır. Genellikle ventral yüzeylerinde seks organları (antheridia ve archegonia) taşıyan kalp şeklinde prothalli (gametofit) üretirler.

Resim Nezaket: upload.wikimedia.org/wikipedia/commons/5/56/Fern_Sori.JPG
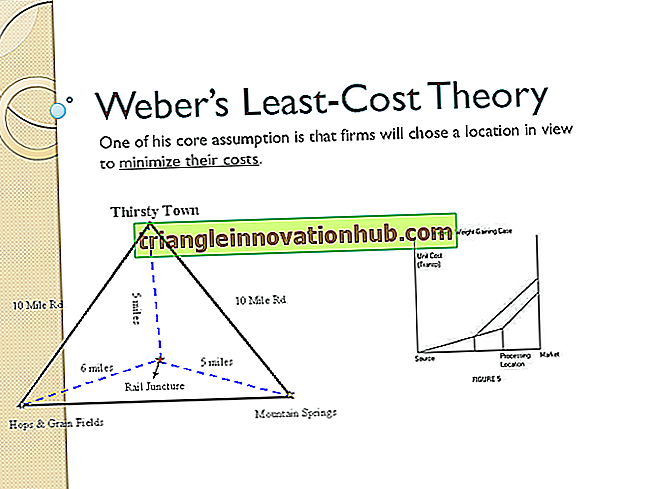
Prothalli, monoecious. Sprangia, sorinde marjinde veya sporofilin yüzeyinde üretilir. Bu düzen, aşağıdaki gibi yedi aileyi içerir:
1. Schizaeaceae
2. Gleicheniaceae
3. Matoniaceae
4. Hymenophyllaceae
5. Dicksoniaceae
6. Cyatheaeae
7. Polypodiaceae

Cins PTERIS Linn:
Sistematik Görev:
Anabilim Dalı: Filicophyta
Sınıf: Leptosporangiopsida
Sipariş: Filicales
Aile: polypodiaceae
Alt Aile: Pteridoideae
Cins: Pteris
Türler: vittata
Yetişme ortamı:
Bu cins, dünyanın tropikal ve alt tropik bölgelerinde bol miktarda yetişen yaklaşık 250 tür tarafından geniş bir şekilde dağılır ve temsil edilir. Yaygın Hint türleri: Pteris vittata, P. cretica, P. biaurita, P. quadriaurita, P. stenophylla ve P. wallichiana. Bununla birlikte, Pteris vittata düşük seviye fem ve yıl boyunca yeni yapraklar ortaya çıkarır.
Dağ duvarları boyunca oldukça yaygındır ve deniz seviyesinden 1200 metreye kadar büyürken, P. quadriaurita kuzey ve batı Himalayalar boyunca vadiler ve vadiler boyunca bol miktarda yetişir. P. cretica, 1200-2400 metre
Alışkanlık:
Bitkiler, P. biaurita, P. vittata ve P. grandiflora'da sürünen rizomlarına sahiptir. P. cretica'da rizom dallı, kısa, sağlam ve yarı dik olarak dallanmıştır. Rizom ölçeklerle kaplıdır. Saç yok. Kökler, köksapın alt yüzeyinden kaynaklanır veya yüzeyin her tarafında ortaya çıkabilir, örneğin, P. biaurita'da (Şekil 31.1).

Yapraklar pinnately bileşiktir. Yaprak sapları pullarla kaplıdır. Yerleşme açık furkate tiptedir. P. vittata'daki yapraklar bir zamanlar pinnattır. Pinnae, tabanın yakınında, ortasının yakınında daha büyük ve yine yaprağın tepesine doğru daha küçüktür. Yaprak tepe eski bir broşür veya pinna tarafından işgal edilmiştir.
Her pinna ucun yakınında çatallanan yan damarları yayan merkezi bir midrib ile geçilir. Pinna tabanda sapsız ve geniştir ve yavaş yavaş tepeye doğru sivrilir, yapraklar P. biaurita'da iki yüzlüdür. Pinnules, coriaceous veya pürüzlüdür (Şekil 31.2).

Anatomi:
Köksapların stelar organizasyonu türden türe, hatta bazen aynı türe bile değişir. P. cretica ve P. vittata'nın genç bölgelerinde basit bir dictyostele olan P. grandiflora ve P. vittata'da solenosteliktir. P. biaurtia'da köksapın genç dallarında stel, alt bölgede karışık bir protosteldir.
Sifonostelik bir parça yükselir ve tepeye yakın solenostelik hale gelir. Ana köksapta da diktiyostelik durum da bulunur. P. vittata'da stelar organizasyonu, köksapın apikal bölgelerinde disiklik bir distiostele dönüşür (Şekil 31.3).
Bu durum, küçük, dairesel veya oval şeritlerin veya kök izlerinin, ana selistellerin dış yüzeyinden ayrılmasıyla sağlanır; bu da solenoz arkın stelar perforasyonu ile elde edilir. Bu teller 3-5 büyük meristel çevresinde düzensiz olarak düzenlenmiş kök izlerinin oluşmasına yol açar (Şekil 31.4). Ancak medüller iplik yoktur.
Pinnule, üst ve alt epidermal katmanlara sahiptir (Şekil 31.5). P. cretica'da üst epidermis, daha az sinüler duvarlı daha büyük hücrelere sahiptir. Stomalar, daha küçük hücrelere ve daha sinirli duvarlara sahip olan alt epidermlerle sınırlıdır.
Mesophyll, palisade ve süngerimsi parankimi olarak ayırt edilemez. Orta kaburga bölgesi, ayrı endodermisli, örneğin P. vittata'da, tek bir eşmerkezli tür vasküler kordona sahiptir. Paket kılıfı uzantıları belirgindir ve alt epidermisin üstünün altında ve üstünde kalın duvarlı hücre grupları olarak ortaya çıkar. Orta kaburga çevresinde palisade ve süngerimsi doku yoktur.

Yaprak sapı, tek bir C-şekilli (P. vittata) veya U-şekilli veya V-şekilli bir yaprak izi ile geçer (Şekil 31.6). P. biaurita'da U-şekilli yaprak izi yaprakçağa girer ve yukarı V-şekilli hale gelir. Ksilem iki adaxial kancasına sahiptir. Ruşilerde, yaprak sapı iz bırakırsa veya yaprak bikinleşirse ayrılmaya devam ederse veya iki yapraklı yapraklarda (P. biaurita) ikincil ve üçüncül pinna izlerine yol açacak şekilde bölündüğü zaman yaprak sapı kılçıklarına ipini verir. Raşis izleri başlangıçta marjinaldir ve genellikle düz U şeklinde veya sığ yay şeklindedir.
Sporofil:
Sorus sürekli tiptedir. Pinnenin kenarlarından oluşan üst indusiyal kapakçık tarafından korunur (Şekil 31.7). Hazne menşeli intramarjinaldir. Düşük veya gerçek indusyum yoktur. Sprangia karışmaz ve sporangia gelişimi Dryopteris ile benzerdir. Her bir sporangium 48 spor üretir.

Sporlar:
Sporlar keskin veya kabaca üçgen şeklindedir ve ayrı bir triradiat maskeye sahiptir. Spor duvarı kalındır ve bir dış omurgaya ve bir iç intine sahiptir. Perispore eksikliği var. Sporlar, türlere göre boyut olarak değişir. Sporun çimlenmesi ve prothallus pozisyonunun oluşması, cinsiyet organlarının dağılımı, yapısı ve gelişimi diğer polipodead üyelerine benzer.
Sporun Çimlenmesi:
Eksine yırtıklar ve uninucleate içerikleri ortaya çıkar ve küçük bir silindirik thallus hücresine dönüşür (Şekil 31.7). Tek hücreli bir rizoid thallus hücresinden ortaya çıkar ve uzun ve tübüler, primer rizoid olarak büyür. Thallus hücresi, yeşil hücrelerin kısa bir filamentini oluşturmak için çapraz olarak bölünür.
Diğer bölümler, enine ve dikey, kalınlıkta bir hücre tabakası tabakası oluşturur. Apikal çentiğin arkasındaki hücreler iki veya daha fazla düzlem boyunca bölünür ve kalın bir merkezi yastık oluşturur. Sekonder rizoitler, prothallun arka bölgelerinden kaynaklanır. Olgun thallus hücreleri, rizoidal ana hücreler olarak görev yapar. Rizoidal hücreler farklılaşır ve ikincil rizoitlere dönüşür.

Cinsiyet organları: anteridia ve archegonia. Cinsiyet organlarının yapısı ve gelişimi Dryopteris'e benzer.
Embriyo ve genç sporofit:
Embriyo ve genç sporofit gelişimi Dryopteris'in gelişimine benzer:
Ekonomik önem:
Pteris türleri çok az ekonomik değere sahiptir. Bunlar, hayvanlara zarar verebilecek zararlı ve toksik yabani otlardır. Bununla birlikte, bazı Pteris türleri yararlıdır. Bitkiler sabun ve cam endüstrisi için bir potas kaynağı olarak kullanılmıştır. Yaprak ve yakıt olarak fronlar kullanılır. Ayrıca tıp ve antelmintik olarak da kullanılırlar. Bitkiler demleme amaçlı kullanılır. Tabak veya boya olarak kullanılırlar. Bazı bitkiler kağıt ve karton yapımında kullanılır.
Cins DRYOPTERIS (Erkek Eğrelti):
Pteridophyta Bölümü:
Filicophyta
Sınıf. Leptosporangiopsida
Sipariş. Filicales
Aile. Polypodiaceae
Genus. Dryopteris
Burada, Polypodiaceae'nin Dryopteris filix-mas'ın (erkek eğrelti otu) ömrünü detaylı olarak tartışacağız.
Eğrelti otları yaşayan pteridofitlerin en bilinenleridir. Erkek eğrelti-Dryopteris filix-mas, hem tepelerde hem de ovalarda gölgeli ve nemli yerlerde bulunur. Genellikle kalın ormanlarda görülür. Bu eğrelti otu çok yaygın ve yaygın bir şekilde dağılmıştır. Hindistan'da, bu ülke genelinde bulunur. Yaygın olarak 2.000 ila 8.000 feet yükseklikteki tepelerde bulunur.
Dış Morfoloji:
Alışkanlık ve kök:
Bu eğrelti otu, yüzeyden yukarı doğru eğimli olarak büyüyen ve yer yüzeyinin çok az üzerinde görünen, kısa, keskin olmayan, sağlam ve dallanmamış bir köksapa sahiptir. Köksap nadiren bir ayak uzunluğundan daha fazla değildir. Dört veya beş inç çapındadır ve tamamen eski yaprakların kalıcı yaprak / bazları ile kaplanmıştır. Köksapın tepesi alt ucundan daha geniştir ve görünüşte konik hale gelir. Bazen, maceraperest tomurcuklar köksap üzerinde de görünür. Bu tomurcuklar yaprak bazlarının hemen üstünde yanal olarak gelişir.

Yapraklar:
Yapraklar ayrıca yaprak olarak da bilinir. Yaprak bileşik ve bir ila üç metre uzunluğundadır. Her bir yaprak çok sayıda broşür veya pinna'dan oluşur. Pinna tekrar küçük piklere bölünmüştür. Yaprak saplı. Yaprak sapı pinnae taşıyan uzun bir rakı ile biter. Genç yapraklarda hem rakı hem de yaprak sapı, ramenta olarak bilinen küçük, kahverengi, tüylü pullarla kaplıdır.
Genç yapraklar sünnetlidir. Yaprak iki yıl içinde olgunlaşır. İlk yıl, yaprak sapı, rakchis ve broşürler kıvrılmış halde kalır. Bu tür genç yapraklara sirkinat yaprakları denir. Olgun yapraklar sadece bir mevsim sürer ve yine köksap üzerinde spiral şekilde düzenlenmiş halde kalan on ila yirmi yaprak çıkar.
Kök:
Kökler birincil ve ikincil olmak üzere iki çeşittir. Embriyo tarafından üretilen birincil kök daha önce ölür. İkincil kökler, aynı zamanda maceracı kökler olarak da adlandırılır. Maceracı kökler, kök dokularından sekonder olarak üretilir. Genellikle her bir sapın tabanında üç maceralı kök gelişir.
Kökler ince ve bol dallıdır, böylece eski bir kök yoğun bir şekilde bir dizi macera köküyle kaplanır. Kökler monopodiyal olarak dallanmıştır, böylece eski bir kök sık sık bir macera kökleri ile kaplanır. Kökler ve dallar acropetal olarak ortaya çıkar.

Dahili Morfoloji:
Köksap anatomisi:
Vasküler silindir tipik bir diktiosteldir. Birçok basit pteridofitte, gövde, korteksle çevrili tek bir merkezi stele sahiptir. Bu eğrelti otunda, diğer birçok eğrelti otunda olduğu gibi, merkezi stel bir halka steline bölünür. Bu steller 'meristeller' olarak bilinir ve stelar sisteminin diktiyostelik olduğu söylenir.
Meristeller, zemin dokusuna düzensiz şekilde gömülür. Dryopteris durumunda, gövde tamamen yaprak tabanlarıyla kaplıdır ve internot yoktur. Sapın herhangi bir enine kesiti, eğik kesik yaprak izlerini gösteren yaprak tabanları ile çevrilidir. Yaprak izleri korteksi geçiyor ve saplara giriyor.
Her meristel, floem ile çevrili merkezi bir ksilem kütlesinden oluşur. Skaleriform tracheids ve ksilem parankimi ksilemi oluşturur. Sap işleminde, tracheids poligonal ve yeterince geniş görünür. Gemiler tamamen yok. Meristele'nin merkezinde, tracheids spiral taşıyor ve kıvrılıyor. Bu tracheids protoksili temsil eder.
Xylem'in geniş tracheid'lerinin geri kalanı metaxylem'i temsil eder. Floem tek bir halka elek tüpünden oluşur. Elek tüpleri oldukça uzundur ve üst üste binen uçlarında sivrilmektedir. İki elek tüpünün birleştiğinde, elek plakaları geliştirilmiştir. Tamamlayıcı hücreler bulunamadı. Elek tüplerinin halkasının hemen altında, iyi miktarda floem parankimi var. Floem, ince duvarlı, tek katmanlı bir pericycle ile çevrilidir.

Meristele hücrelerinin en dış tabakasıdır. Her meristel bir endodermis ile çevrilidir. Endodermis, korteksin son tabakasıdır. Casparian bantları endodermal hücrelerin radyal duvarlarında bulunur. Endodermisin dışında, epidermise kadar uzanan kortikal parankimi bulunur.
Meristel, eşmerkezli ve amfibidir. Dryopteris'te meristeller oval ve bağımsızdır. Diktiostelin tamamını saran ortak bir endoderm yoktur.
Korteks, meristellerin gömüldüğü bir kit parankimiden (zemin dokusu) oluşur. En korteks hücrelerinin birkaç katmanı sklerenkimatözdür. Kökün en dış tabakası, kalın dış duvarlara sahip hücrelerden oluşan epidermistir. Epidermis üzerinde bir hücreli kalın ramenta bulunur. Bu skala benzeri ramentanın modifiye edilmiş kıllar olduğu düşünülmektedir.
Yaprak anatomisi:
Anatomi açısından, yaprak (a) yaprak sapı ve (b) lamina olmak üzere iki kısma ayrılabilir.
(a) Yaprak sapı:
Yaprak sapının anatomisini incelemek için birinin enine kesitlerini kesmesi gerekir. Yaprak sapı anahat olarak yaklaşık yarım daire şeklindedir. Yaprak sapının üst kısmı (adaxial) düzdür ve gövdeye doğru durmaktadır. En dıştaki katman epidermistir. Epidermisin hemen altında birkaç katman sklerenkima hücresi bulunur.
Meristeller parankime gömülüdür (zemin dokusu). Düzenlemede meristeles at nalı gibidir. Raşilerin adaksiyel tarafında, her biri her bir pinnaya dallar veren iki meristel vardır.

(b) Lamina:
Lamina bir üst ve alt epidermise sahiptir. Üst epidermisin hemen altında, aralarında iyi gelişmiş hücreler arası boşluklara sahip olan düzensiz parankimatöz hücrelerden oluşan mezofil vardır. Hem epidermal hem de mezofil hücreleri kloroplast içerir. Epidermisteki kloroplastın varlığı, gölgeyi seven bir bitkinin özelliğidir.
Laminanın alt tarafında alt epidermis bulunur. Alt epidermisin hemen altında, aralarında geniş hücreler arası boşluklara sahip düzensiz dallanmış ve yıldız şeklindeki hücrelerden oluşan mezofil vardır. Vasküler teller konsantriktir.
Kök anatomisi:
Kökün anatomisini incelemek için, maceracı kökün enine kesitleri gerekir. Kökün iç yapısı oldukça basittir. Floem ile çevrili merkezi bir ksilem merkezi vardır. Ksilem iki ucunda yer alan iki protoksil grubuyla birlikte. Bu şekilde, ksilem ishal olur. Ksilem dışında floem elementleri vardır.
Floem, floem plakaları şeklinde, ksilime yanal olarak düzenlenir. İki katmanlı bir pericycle, floemi çevreliyor. Pericycle stelin en dış kısmıdır. Daha sonra stelin etrafında tek katmanlı bir endoderm bulunur.
Casparian şeritleri endodermisteki hücrelerin radyal duvarlarında bulunur. Kökün enine bölümünün en dış tabakası, pilifer tabakasıdır. Kazıklı tabaka tek tabakalıdır ve üzerinde ve orada kök kılları taşır. Kortikal bölge iki bölgeye ayrılır. Dış bölge, ince cidarlı hücrelerden, iç kısmı ise kalın cidarlı hücrelerden oluşmaktadır.
Apikal büyüme:
Tüm eğrelti otlarında, saplar, tek apikal hücre gibi üç taraflı bir piramitten büyür. Apikal hücre, olgun dokunun hücrelerini geliştiren üç tarafının her birine paralel olarak tekrar tekrar bölünür.
Kökler ayrıca apikal hücreye benzer üç taraflı bir piramitten de gelişir. Hücrenin bu üç tarafından kökün bütün organları tekrarlanan bölünmeleri ile gelişir. Bununla birlikte, kök başlığının hücreleri, apikal hücrenin tabanından kesilir.
Spor üreten organlar:
Sprangia, sporofiller olarak bilinen yapraklar üzerine yerleştirilmiş sorilerde üretilir. Sori, yaprakların kepeğinin alt yüzeyinde bulunur. Dryopteris'te vejetatif yapraklar ve sporofiller birbirine çok benzer, ancak sporofiller eski bitkilerde görülür. Sprangia'nın küçük böbrek şeklindeki sorisi, yaprağın piklerinin alt tarafında iki sıra halinde düzenlenmiştir.
Bir sorusun sporangiaları şişmiş bir plasentaya saplarıyla tutturulur. İndusyum olarak bilinen hassas bir membranöz yapı, plasentadan da ortaya çıkar ve sporangia sorusunu bir örtü gibi kaplar. Olgun sporangia rengi kahverengidir.

Sporangium gelişimi:
Sporangium gelişimi diğer Polipodiaceae cinslerinde olduğu gibi leptosporangiat tipindedir. Sprangium, plasentanın tek bir yüzeysel hücresinden gelişir. Bu hücre, iki hücre oluşturan enine bir duvarla bölünür. Üst hücre yine eğik bir duvarla ilk duvara bölünür.
Tetrahedral bir hücreye yol açan başka bir eğik bölünme gerçekleşir. Bu hücre yine enine bir duvarla bölünür. İç hücre, kemirgen hücresi olarak bilinir. Birinci bölünme tarafından oluşturulan çiftin alt hücresi birkaç kez bölünür ve çok hücreli sapa doğru gelişir. Tetrahedral archesporial hücre çevreleyen hücreler ayrıca sporangial duvarına yol açan bölünürler.
Bundan sonra, kemerler arası hücre, hücrelerin bir kaset tabakası oluşturmak için bölünür. Aynı zamanda yaklaşık bir düzine spor anne hücresi üretir. Spor anne hücreleri sporlara yol açar. Her spor anne hücresi iki kez gelişerek dört sporu böler. Bölünme sırasında mayoz var ve sporlar haploid. Genellikle her bir sporangium 48 haploid spor içerir.
Sporangium yapısı:
Her bir sporangium çok hücreli bir sap ve bir kapsülden oluşur. Sprangium'nun kapsülü, bir bikonveks lens şeklindedir. Kapsülün duvarı, tek bir ince duvarlı hücre katmanından oluşur. Halka olarak bilinen özel olarak kalınlaşmış hücrelerden oluşan bir sıra, kapsülü kısmen çevreler. Annülüsün bir tarafında, ince cidarlı hücrelerden oluşan bir mide vardır. Olgunlukta, sporangium içinde 32-64 koyu, düzensiz duvarlı spor içerir.

Sporangium'nın ıslahı:
Sprangia'nın olgunlaşması üzerine, indusyum kırışır ve sporangia dış atmosfere maruz kalır. Sprangium özel bir mekanizma ile patlar. Halka şeklindeki hücreler tuhaf bir şekilde kalınlaşır. Halka şeklinin iç teğetsel ve radyal duvarları kalın iken dış teğetsel duvarlar incedir.
Sprangium'nun patlaması, suyun annulus hücrelerine göre davranışlarından kaynaklanır. Genç sporangia'da su bezleri bulunur ve nemli kalırlar. Sprangia'nın olgunluğunda, sporangia kuru havaya maruz bırakılır ve sporangia hücrelerinden su kaybolur.
Bu şekilde, halkanın ince dış duvarları büzülmeye başlar, mide açılır ve sporangium'nun bir kısmı ile birlikte annulus, bir sperk ile geriye doğru döner ve sporları sporangium'dan dışarı atar.

Spor:
Sporlar çift duvarlıdır. Dış duvar kalın, düzensiz ve kahverengi renktedir. Eksine olarak bilinir. İç duvar pürüzsüz ve incedir ve intine olarak bilinir. Her sporda büyük bir merkezi çekirdek vardır. Feminin gametofitik evresi bu spordan başlar.
Sporun Çimlenmesi ve Protestusun Gelişimi:
Dağılımlarından sonra sporlar bir süre uykuda kalırlar ve uygun nem ve sıcaklık koşulları altında filizlenirler. Öncelikle dış spor duvarı (exine) kırılır ve daha sonra intine yeşil germ tüpü şeklinde dışarı çıkar. Çok kısa bir süre içinde bu tüp enine bir duvar tarafından kesilir ve iki eşit olmayan hücre ortaya çıkar. Küçük renksiz hücre, rizoidal hücre olarak bilinir. Bu hücreden renksiz bir dal toprağa doğru büyür ve bir rizoid geliştirir. Büyük yeşil hücre uzar ve bölünür

enlemesine birkaç kez uzunluğu 3 ila 6 hücreye sahip bir filament haline gelir. Apikal bir hücre olarak bilinen filamentin terminal hücresi, arkasındaki hücreleri keser ve bu filamentli yapıya yol açar. Apikal hücre ayrıca yanal olarak bölünür ve düz prothallus oluşur. Apikal hücreden kesip kesilen parçalar tekrar tekrar bölünerek genç proteale dönüşür. Genellikle prothallus kalp şeklindedir.
Prothallus yeşildir ve kenarlarından birinde kalın bir hücre vardır. İyi gelişmiş prothallide, prothallusun merkezi kısmı, kalınlıkta birden fazla hücredir. Bu bölgeye yastık denir. Prothallusu toprağa sabitleyen yastığın alt kısmında birçok rizoit gelişir. Rizodiler ayrıca emici organlar olarak da işlev görür.
Gametofitin gelişimi sırasında orijinal apikal hücre, bir grup marjinal baş harfle değiştirilir. Protüsün daha da büyümesi bu baş harflerle gerçekleşir ve sonuçta kalp şeklindeki prothall oluşur.
Prothallusun yapısı:
Prothallus düz, kalp şeklinde ve yeşil bir doku kütlesidir. Arka ucunda bir çentik var. Kenarlarında kalınlıkta bir hücredir, ancak merkezi bölgede, çentik yakınında, pek çok kalınlıkta hücrelidir. Bu bölgeye yastık denir. Yastığın altından birkaç renksiz rizoit gelişmiştir.
Prothallusun hücreleri ince duvarlıdır. Her bir hücre diskoid kloroplast ve içinde bir çekirdek taşır. Cinsiyet organları (archegonia ve antheridia), prothallun orta bölgesinde sınırlıdır. Her prothall, muhtemelen 1/8 ila 1/3 inç çapındadır. Gırtlak kendini destekliyor. Topraktaki suyu ve besinleri emer ve hücrelerinde çok sayıda kloroplast bulunduğundan besin materyalini fotosentez eder.

Prothallus, üzerinde seks organlarını taşıdığı için eğrelti otunun gametofitik neslini temsil eder.

Cinsiyet organları:
Seks organları, prothallusun (gametofit) alt kısmında gelişir. Genellikle prothalli monokülerdir ve hem erkek hem de kadın cinsiyet organlarını, yani anteridia ve archegonia'yı taşır. Normal prothallda archegonia, yastık üzerindeki çentik çevresinde ve prothallusun bazal merkezi bölgesinde rizoidler arasında antantezi oluşturur. Anteridia ilk önce archegonia gelişir.

Anteridyumun gelişimi:
Prothallusun yüzeysel vejetatif hücrelerinden biri, iki hücreye yol açan enlemesine bir duvarla dışarı çıkmakta ve ayrılmaktadır. Terminal hücre, anteridial bir başlangıç görevi görür. Öncelikle anteridiyal başlangıç, geniş ucu yukarı bakacak şekilde eğik huni şeklinde bir duvarla bölünür. Ardışık böyle bir eğik bölünme ile merkezi bir hücre oluşur.
Bu merkezi hücre, orijinal başlangıçta kendilerinde kloroplastları içeren birkaç çevresel hücre ile çevrilidir. İlk bölünmeyi, huninin tepesi boyunca kubbe şeklinde bir duvar izler ve böylece merkezi bir hücre içine alınır. Bundan sonra, üst hücre halka şeklindeki bir duvarla bölünür.
Bu şekilde, yarı küresel anteridyum iki halka hücreden, bir kap hücreden ve bir merkez hücreden oluşur. Merkezi hücre düzensiz olarak bölünür ve antherozoid ana hücreler (andrositler) olarak bilinen yaklaşık otuz iki hücre üretilir. Her bir antherozoit ana hücre (androsit), bir multiciliate antherozoide metamorfozlar.
Her andrositin çekirdeği, üç turlu bir konik spiral haline dönüştürülür. Bu sarmal yapının arka ucuna, bir sitoplazma keseciği eklenir. Ön ucunda, çok uzun bir flagella grubunu taşıyan uzun bir blefaroplast var. Flagella vasıtasıyla antherozoitler su filminde yüzebilirler.

Anteridyumun alçalması:
Su bu işlem için çok önemlidir. Olgun anteridyum suyla temas ettiğinde, olgun anteridyumda bulunan andrositlerin duvarları yapışkan hale gelir ve şişer ve sonuçta anteridyumun kapak hücresi atılır. Antherozoitler dışarı atılır. Her bir antherozoit, ince bir membranöz hücre çeperi içindedir, çok yakında suda çözülür ve anterozoid suda serbest kalır.
Archegonium'un gelişimi:
Archegonium, archegonial başlangıç olarak bilinen, prothallusun tek bir yüzeysel hücresinden gelişir. İlk, iki enine duvar tarafından üç hücreye ayrılır. Bu üç hücrenin alt tarafı, venter hücresi etrafındaki duvarı oluşturan bazal hücredir. Kalan iki hücre, merkezi hücre ve örtü hücresidir.
Kapak hücresi dört boyunlu baş harfleri oluşturan dikey olarak iki kere bölünür. Her boyun başlangıcı, enine gelişerek, boyun hücrelerinin dört dikey sırasını böler. Boyun 5 ila 7 hücrelidir. Merkez hücre, boyun hücreleri arasında büyür.
Bu, birkaç kez enine şekilde bir dar boyunlu kanal hücresi, bir ventral kanal hücresi ve bir yumurta veya oosfer üreterek böler. Boyun kanalı hücresi, iki hücreye bölünebilir veya olmayabilir. Boyun keskin bir şekilde kavisli. Dışbükey tarafın boynundaki iki sıra 5 ila 7 hücre içerir, oysa içbükey tarafın sıraları sadece dört hücreye sahiptir ve dolayısıyla boyun kuvvetli bir şekilde kavislidir.
Archegonium'un yapısı:
Archegonium, proteal dokularına gömülü bir delikten ve proteal dokularının dışına çıkıntı yapan keskin bir kavisli boyundan oluşan, şişeye benzeyen uzun bir yapıdır. Kavisli boyun dört dikey hücre sırasından oluşur. Boynundaki ve venterindeki archegonium, bir eksenel hücre sırası içerir.
Kanalın içinde bulunan en alt hücre oosfer veya ovumdur; Üstünde bir ventral kanal hücresi vardır ve bundan sonra genellikle binüklüt olan uzun boyunlu bir kanal vardır. Döllenmeden önce, boyun kanalı hücresi ve ventral kanal hücresi, müsilajinli bir maddeye dönüşür. Bu yapışkan malzeme şişer ve boynun dışına doğru zorlar. Bu şekilde, boynun tepesinde bir açıklığa neden olur ve anterozoitler bu açıklıktan oosfere ulaşmak için girer.

Döllenme:
Su bu işlem için önemli bir faktördür. Normalde prothalli yağmurla su alır. Olgun archeg-onium muhtemelen yüzme antherozoitlerini kendine çeken malik asidi salgılar. Antheroz-oids boyun kanalından musilaj boyunca yüzüyor ve bunlardan biri oosferde hiyalin alıcı noktasında nüfuz ediyor.
Antherozoidin yumurtaya girmesinden kısa bir süre sonra iki erkek ve dişi çekirdek birbirine kaynaşır ve döllenme gerçekleşir. Ososfer etrafındaki bir duvarı salgılar ve oospore olur. Tek bir prothallun birkaç argonyası döllenebilir, ancak sonuçta sadece bir oospor tam sporofit içinde gelişir.

Embriyo ve gelişimi:
Döllenmeden hemen sonra zigot iki hücre oluşturan dikey bir duvarla bölünür. Kısa bir süre sonra ikinci ve üçüncü bölünmeler birbirleriyle dik açılarla meydana gelir ve bir oktan veya sekiz hücre grubuyla sonuçlanır. Ana prothallusun zirvesine doğru bulunan oktan hücrelerinin dördü, epibazal yarımı ve kalan dört hücreyi hipobasal yarısı oluşturur.
Epibasal hücreler, ekseni ve genç sporofitin ilk yaprağını üretir. Hipobasal yarısı primer kökü ve ayağı üretir. Ayak, haustoryum gibi davranır ve karın içine gömülü kalır. Genç sporofitin, prothallustan ve topraktan bağımsız olup olmadığı sürece beslenmeyi emer.
Birincil kök yakında ölür ve maceracı kökler gelişir. Prothallus çok yakında dağılır. İlk yaprak yiyeceği sentezleyebildiği anda prothallus kaybolur. Genç sporofitin ilk yaprağı genellikle küçük ve iki lobludur. Kısa süre sonra iki büyük çatal biçimindeki daha büyük yapraklar gelişir. Sadece beşinci ve altıncı yaprakların oluşumundan sonra, iğneli bileşik yaprakları gelişir.

Nesiller Alternatif:
Kromozom sayısı iki katına çıkar, yani döllenme bittikten sonra diploid (2n). Bu diploid evre, spor anne hücrelerinin oluşumuna baskın gelir. Bu aşamada mayotik bölünme meydana gelir ve haploid («) sporları oluşur. Sporlar, gametofitik aşamanın yeni başlayanlarıdır. Spor çimlenir ve prothallus oluşur.
Anteridia ve archegonia, prothallusta gelişir. Antherozoid ve ov kaynaşmış ve döllenme gerçekleşmiştir. Zigot, sporofitik evrenin başlangıcıdır. Sporofit (2ri), gametofit (prothallus) 'a neden olur ve yine gametlerin birleşmesiyle sporofitik aşamaya ulaşılır. Bu şekilde, eğrelti otunun bir haploid (gametofitik) ve diploid (sporofitik) kuşak alternatifi gösterdiği söylenebilir.
Cins ADIANTUM:
Sistematik pozisyon Pteridophyta
Bölünme. Filicophyta
Sınıf. Leptosporangiopsida
Sipariş. Filicales
Aile. Polypodiaceae
Genus. adiantum
Bower'a göre bu cinste yaklaşık 184 eğrelti otu vardır.
Bu cinsin türü, dünyanın tropikal bölgelerinde yaygın olarak dağılmıştır. Gölgeli ve nemli ortamlarda bulunurlar. Ülkemizde türler Himalayaların engebeli bölgelerinde ve ovalarda oldukça yaygındır.
Alışkanlık:
Adiantum türleri dik veya sürünen pullu bir köksapa sahiptir. Yapraklar rizom üzerinde sarmal olarak veya dönüşümlü olarak düzenlenmiştir. Yaprak sapı siyah ve kırılgan parlıyor. Bıçak, A. reniforme'de tamamen olabilir veya basit veya tekrar tekrar dallanmış olabilir. Bıçaklar dokularında oldukça hassastır.
Çatallı damarlar bıçağı çaprazlar. Genellikle damarlar serbesttir, ancak bazı türlerde retikülü oluşturan anastamossajlar vardır. Yaprağın dallanması iki türdendir. Nihai bölümler anahat olarak deltoid edilir. Eğrelti kızlık saç eğreltiotu olarak bilinir.

Köksap anatomisi:
Uzatılmış bir köksapa sahip olan Adiantum türleri, örneğin A. pedatum'da gerçek solenostelleri gösterebilir, ancak genel olarak stel, yaprak boşluklarının uzaması nedeniyle oluk şeklindedir. Kısa iç içe sahip olan ve köksapın dik olduğu bu köksaplarda, yaprak boşlukları üst üste binmekte ve stelin enine bölümünde görüldüğü gibi bir halka içinde düzenlenmiş birkaç meristel bulunmaktadır.
Yaprak anatomisi:
Yaprak tabanı, tek bir dört açılı meristel oluşturmak için yukarı doğru birleşen birbirine yakın iki ipi alır. 'Spicullar hücreler' Adiantum bıçağının epidermal hücrelerinde bulunmuştur.
Kök anatomisi:
Kökün anatomisini incelemek için, maceracı kökün enine kesitleri gerekir. Kökün iç yapısı oldukça basittir. Floem ile çevrili merkezi bir ksilem merkezi vardır. Ksilem iki ucunda yer alan iki protoksil grubuyla birlikte. Bu şekilde, ksilem ishal olur. Ksilem dışında floem elementleri vardır. Floem, floem plakaları şeklinde, ksilime yanal olarak düzenlenir.
İki katmanlı bir pericycle, floemi çevreliyor. Pericycle stelin en dış kısmıdır. Daha sonra stelin etrafında tek katmanlı bir endoderm bulunur. Casparian şeritleri endodermisteki hücrelerin radyal duvarlarında bulunur.
Kökün enine bölümünün en dış tabakası, pilifer tabakasıdır. Kazıklı tabaka tek tabakalıdır ve üzerinde ve orada kök kılları taşır. Kortikal bölge iki bölgeye ayrılır. Dış bölge, ince cidarlı hücrelerden, iç kısmı ise kalın cidarlı hücrelerden oluşmaktadır.
Apikal büyüme:
Tüm eğrelti otlarında, saplar, tek apikal hücre gibi üç taraflı bir piramitten büyür. Apikal hücre, olgun dokuların hücrelerini geliştiren üç tarafının her birine paralel olarak tekrar tekrar bölünür.
Kökler ayrıca apikal hücreye benzer üç taraflı bir piramitten de gelişir. Hücrenin bu üç tarafından, kökün tüm organları tekrarlanan bölünmelerle gelişir. Bununla birlikte, kök başlığının hücreleri, apikal hücrenin tabanından kesilir.
Spor üreten organlar:
Adiantum'un karakteristik özelliği, kökeni yüzeysel olan ve olgunlaşan zar rengi ve kahverengi renginden dolayı irisyum gibi görünen keskin refleksli bir yaprak marjı ile kaplanmış, görünüşte marjinal sorinin varlığıdır. Aslında gerçek bir indusyum yok.
Sprangrangia, fertil lobu geçen damarların distal bölgesine yerleştirilir. Bıçağın bereketli bölgesi keskin bir şekilde geri yansıtılır ve indusyum görevi görür. Bu bereketli lobların her biri, paralel damarların üzerine yerleştirilmiş bir grup sori taşır. Bazı durumlarda, sporangia bıçağın yüzeyinde damarlar arasında yayılır.

Sporangium gelişimi:
Sporangium gelişimi, diğer tüm Polypodiaceae cinslerinde olduğu gibi cüzzam türündedir. Sprangium, plasentanın tek bir yüzeysel hücresinden gelişir. Bu hücre, iki hücre oluşturan enine bir duvarla bölünür.
Üst hücre yine eğik bir duvarla ilk duvara bölünür. Tetrahedral bir hücreye yol açan başka bir eğik bölünme gerçekleşir. Hücre tekrar enine bir duvarla bölünür. İç hücre, kemirgen hücresi olarak bilinir. Birinci bölünme tarafından oluşturulan çiftin alt hücresi birkaç kez bölünür ve çok hücreli sapa doğru gelişir.
Tetrahedral archesporial hücre çevreleyen hücreler ayrıca sporangial duvarına yol açan bölünürler. Bundan sonra, kemerler arası hücre, hücrelerin bir kaset tabakası oluşturmak için bölünür. Aynı zamanda yaklaşık bir düzine spor anne hücresi üretir. Spor anne hücreleri sporlara yol açar. Her spor anne hücresi iki kez gelişerek dört sporu böler. Bölünme sırasında mayoz var ve sporlar haploid. Genellikle her bir sporangium 48 haploid spor içerir. (Ayrıca bkz. Şekil 31.6).
Sporangium yapısı:
Her bir sporangium çok hücreli bir sap ve bir kapsülden oluşur. Bu sporangium'nun kapsülü bikonveks lens şeklindedir. Kapsülün duvarı, tek bir ince duvarlı hücre katmanından oluşur. A row of specially thickened cells, known as annulus partially surrounds the capsule. At one side of the annulus, there is a stomium composed of thin walled cells. On maturity, the sporangium contains about 32 to 64 dark, ragged walled spores in it.
Dehiscence of sporangium:
On the maturity of the sporangia, indusium shrivels up and the sporangia are exposed to the outer atmosphere. The sporangium bursts by a special mechanism. The cells of the annulus are thickened in a peculiar way. The inner tangential and radial walls of the annulus are thick while the outer tangential walls are thin.
The bursting of the sporangium is caused by the behaviour of the water in relation to the cells of the annulus. In young sporangia the water glands are present and they remain moist. On the maturity of the sporangia, the sporangia, are exposed to dry air and the water is lost from the cells of the sporangia. This way, the thin outer walls of the annulus begin to contract, the stomium opens and the annulus along with some portion of the sporangium turns backward with a jerk throwing the spores out of the sporangium.
Spor:
The spores are double walled. The outer wall is thick, ragged and brown in colour. It is known as exine. The inner wall is smooth and thin, and known as intine. Each spore contains a big central nucleus. The gametophytic stage of the fern begins from this spore.
Germination of the spore and development of the prothallus:
After their dispersal the spores lie dormant for some time and under favourable conditions of moisture and temperature, they germinate. First of all outer spore wall (exine) breaks and thereafter the intine protrudes out in the form of a green germ tube. Very shortly this tube is cut off by a transverse wall and two unequal cells are resulted.
The small colourless cell is known as rhizoidal cell. From this cell a colourless branch grows into the soil and develops a rhizoid. The large green cell elongates and divides transversely several times developing into a filament 3 to 6 cells in length.
The terminal cell of the filament known as an apical cell cut of cells behind it and gives rise to this filamentous structure. The apical cell also divides laterally and a flat prothallus is formed. The segments cut off from the apical cell divide again and again developing into the young prothallus. Usually the prothallus is heart-shaped.
The prothallus is green and one-celled thick at its margins. In well developed prothalli the central part of the prothallus is more than one cell in thickness. This region is called cushion. Many rhizoids develop on the underside of the cushion anchoring the prothallus to the soil. The rhizoids also serve as absorbing organs.

During the development of the gametophyte the original apical cell is replaced by a group of marginal initials. The further growth of the prothallus takes place by these initials and ultimately the heart-shaped prothallus is formed.

Structure of prothallus:
The prothallus is flat, thin, heart-shaped and green mass of tissue. There is a notch at its posterior end. It is one cell in thickness at its margins, but in the central region, near about the notch, it is many celled in thickness. This region is called the cushion. Several colourless rhizoids are developed from the underside of the cushion.
The cells of the prothallus are thin walled. Each cell bears discoid chloroplasts and a nucleus in it. The sex organs (archegonia and antheridia) are confined in the central region of the prothallus. The prothallus is probably 1/8 to 1/3 of an inch diameter.
Each prothallus is self supporting. It absorbs water and nutrients from the soil and photosynthesizes the food material as the numerous chloroplasts are found in its cells. The prothallus represents the gametophytic generation of the fern as it bears the sex organs upon it.
The sex organs:
The sex organs develop on the underside of the prothallus (gametophyte). Usually the prothalli are monoecious and bear both male and female sex organs, ie, antheridia and archegonia. In the normal prothallus the archegonia develop around the notch upon the cushion and the antheridia in the basal central region of the prothallus among the rhizoids. The antheridia develop first and the archegonia later on.

Anteridyumun gelişimi:
One of the superficial vegetative cells of the prothallus protrudes out and divides by a transverse wall giving rise to two cells. The terminal cell acts as an antheridial initial. First of all the antheridial initial divides by an oblique funnel- shaped wall with the wide end upwards. By successive such oblique divisions a central cell is formed.
This central cell is surrounded by a number of peripheral cells containing the chloroplasts in them of the original initial. The first division is followed by a dome shaped wall across the top of the funnel and thus a central cell is enclosed. Thereafter, the upper cell divides by a ring-shaped wall.
This way, the semispherical antheridium is composed of two ring cells, a cap cell and a central cell. The central cell divides further irregularly, and about thirty two cells known as antherozoid mother cells (androcytes) are produced. Each antherozoid mother cell (androcyte) metamorphoses into a multiciliate antherozoid.

The nucleus of each androcyte is transformed into a tapering spiral of three turns. At the posterior end of this spiral structure a vesicle of cytoplasm is attached. At the anterior end there is an elongated blepharoplast bearing a group of very long flagella. By means of flagella the antherozoids can swim in the water firm.

Dehiscence of antheridium:
The water is essential for this process. When the mature antheridium comes in contact with the water, the walls of the androcytes situated within the mature antheridium become mucilaginous and swell and with the result the lid cell of the antheridium is thrown off. The antherozoids are ejected out. Each antherozoid is still within a thin membranous cell wall, which very soon dissolves in the water and the antherozoid is free in the water.

Archegonium'un gelişimi:
The archegonium develops from a single superficial cell of the prothallus, known as the archegonial initial. The initial divides into three cells by two transverse walls. Of these three cells the lower one is the basal cell which forms the wall around the venter. The remaining two cells are the central cell and the cover cell.
The cover cell divides vertically twice forming four neck initials. Each neck initial divides transversely developing four vertical rows of the cells of the neck. The neck is 5 to 7 cells in height. The central cell grows up between the neck cells.
This divides several times transversely producing a narrow neck canal cell, a ventral canal cell and an egg or oosphere. The neck canal cell may or may not divide further into two cells. The neck is sharply curved. The two rows of the neck of the convex side contain 5 to 7 cells, whereas the rows of concave side have only four cells and thus the neck is strongly curved.


Archegonium'un yapısı:
The archegonium is flask-like, elongated structure consists of a venter embedded in the tissues of the prothallus and a sharply curved neck which projects outside the tissues of the prothallus. The curved neck consists of four vertical rows of cells. The archegonium with its neck and venter contains an axial row of cells.
The lowermost cell found within the venter is the oosphere or ovum, above to it there is a ventral canal cell and thereafter a long neck canal cell which is generally binucleate. Prior to fertilization the neck canal cell and the ventral canal cell break down into a mucilaginous substance. This mucilaginous material swells and forces its way outside the neck. This way an opening is caused at the apex of the neck and the antherozoids enter through this opening to reach the oosphere.
Döllenme:
The water is an essential factor for this process. Ordinarily the prothalli receive water by rain. The mature archegonium probably secretes malic acid, which attracts the swimming antherozoids towards it. The antherozoids swim down the neck canal through the mucilage and one of those penetrates the oosphere at a hyaline receptive spot.
Soon after the entrance of the antherozoid in the egg the two male and female nuclei are fused together and the fertilization is effected. The oosphere secretes a wall around it and becomes oospore. Several archegonia of a single prothallus may be fertilized but ultimately only one oospore develops into the complete sporophyte.

The embryo and its development:
Immediately after fertilization the zygote divides by a vertical wall, forming two cells. Soon after second and third divisions take place at right angles to each other resulting in an octant or a group of eight cells. Four of the cells of the octant found

towards the apex of the prothallus form the epibasal half and the remaining four cells the hypobasal half. The epibasal cells produce the axis and the first leaf of the young sporophyte. The hypobasal

half produces the primary root and the foot. The foot acts as haustorium and remains embedded in the venter. It absorbs nourishment for the young sporophyte from the prothallus and soil unless and until it becomes independent. The primary root dies soon and the adventitious roots develop. The prothallus disintegrates very soon. As soon as the first leaf becomes able to synthesize the food, the prothallus disappears. The first leaf of the young sporophyte is generally small and two- lobed. Soon after other larger leaves develop which are dichotomously forked. Only after the formation of the fifth and sixth leaves, the pinnately compound leaves are developed.

Alternation of generations:
The number of chromosomes becomes doubled, ie, diploid (2n) after fertilization is over. This diploid stage prevails upto the formation of spore mother cells. Meiotic division takes place at this stage and the haploid (n) spores are formed.
The spores are the beginners of the gametophytic stage. The spore germinates and prothallus is formed. The antheridia and archegonia develop on the prothallus. The antherozoid and ovum fuse and fertilization is effected.
The zygote is the beginning of the sporophytic stage. The sporophyte (2n) gives rise to gametophyte (prothallus) and again by the process of the union of the gametes the sporophytic stage is attained. This way, it may be said that the fern exhibits an alternation of haploid (gametophytic) and diploid (sporophytic) generations.