Golgi Kompleksi: Terminoloji, Oluşum, Morfoloji, Yapı ve Diğer Detaylar
Golgi Kompleksi: Terminoloji, Oluşum, Morfoloji, Yapı, Yapısal ve Biyokimyasal Polarizasyon, Kimyasal Bileşimi, İşlevleri ve Kökenleri!
1898 yılında gümüş boyama yöntemiyle Golgi, sitoplazmada retiküler bir yapı keşfetti. Genellikle bu yapıya verilen “Golgi aparatı” ismi kafa karıştırıcıdır çünkü hücrenin fizyolojik süreçleri ile kesin bir ilişki olduğunu gösterir.
Bugün, özel boyama özelliklerine sahip olan bu malzemeye atıf yapmak için “Golgi özü” veya “Golgi kompleksi” adını kullanmak daha uygun görünmektedir. Kırılma indisi matrikse benzer olduğundan Golgi kompleksinin canlı hücrelerde gözlemlenmesi zordur. Elektron mikroskobunun kullanımı, bu bileşenin farklı bir görüntüsünü sağlamıştır ve mikroskobik alt yapısı ortaya çıkarılmıştır.
Yıllarca Golgi kompleksinin çeşitli fiksasyon ve boyama prosedürlerinin bir eseri olduğu düşünülüyordu. Başka bir deyişle, birçok bilim adamı sayısız mikroskopi prosedürü sırasında gözlemlenen ve Golgi olarak adlandırılan yapının canlı hücrede bulunmadığına inanmaktadır. Guilliermond (1923), Parat (1927), Walker ve A lien (1921), Golgi kompleksinin varlığından kuşku duyuyordu.
Golgi kompleksinin terminolojisi:
Holmgren Golgi kompleksine trophospongium olarak anılır (Cajal, Golgi-Holmgren kanalları olarak anılır). Baker, varsayılan lipit içeriği nedeniyle lipokondri terimini kullandı. Dalton Kompleksi terimi, 1952'de gözlemcisi Dalton adından sonra verildi. Sjostrand, Golgi sistemi için sito-membranlar terimini önerdi.
Sosa, Golgi kompleksi için aşağıdaki isimlendirmeyi önermiştir:
1. Golgiokinesis :
Golgi aparatının nükleer bölünme sırasında bölünmesi.
2. Golgiyomlar :
Golgiogenesis tarafından üretilen organlara omurgasızlarda Golgi materyali olarak tanımlanan Golgiosomlar adı verilir.
3. Golgiolysis :
Golgi aparatının çözünme süreci.
4. Gogiorreksis :
Golgi cihazında parçalanma.
5. Golegiogenez :
Embriyonik gelişim sırasında Golgi vücudunun oluşumu ve farklılaşması.
6. Golgio-cytoarchitecture :
Golgi aparatına göre hücre yapısının incelenmesi.
Oluşum:
Golgi kompleksi prokaryotik hücreler (viz., Mycoplasma, bakteri ve mavi yeşil algler) ve bazı mantarların ökaryotik hücreleri, briyofitler ve pteridofitlerin sperm hücreleri, olgun elek tüplerinin ve olgun sperm ve kırmızı kan hücrelerinin sperm hücreleridir. hayvanların hücreleri.
Golgi kompleksinin morfolojisi:
Golgi kompleksinin morfolojisi, bulundukları hücrenin türüne bağlı olarak hücreden hücreye değişir. İki Golgi kompleksi formu gözlenmiştir.

1. Yerelleştirilmiş form :
Omurgalıların kutuplaşmış hücrelerinde (baz ve tepe), Golgi kompleksi tek başına oluşur ve sabit bir pozisyonda bulunur. Çekirdek ile salgı direği arasında uzanır. Bu en iyi tiroid hücrelerinde, ekzokrin pankreas hücrelerinde ve bağırsak mukozasında görülebilir.
2. Dağınık form :
Bazı özel omurgalı hücrelerinde (sinir hücreleri ve karaciğer hücreleri), çoğu bitki hücresinde ve omurgasız hücrelerinde, birkaç ünite Golgi kompleksi, endoplazmik retikulum elementleri ile birlikte dağılmış olarak bulunur. Her birime bir sözlük olarak adlandırılır. Karaciğer hücrelerinde, hücre başına 50 kadar diktiyom vardır ve bazı bitki hücrelerinde sayıları yüzlerce sayıya ulaşabilir.
Şekil :
Golgi kompleksinin şekli, farklı somatik hücre türlerinde hayvanlarda oldukça değişkendir. Aynı hücrede bile farklı fonksiyonel aşamalarda farklılıklar vardır. Bununla birlikte, şekil her hücre tipinde sabittir. Kompakt bir kütleden dağılmış bir filamentli ağa kadar çeşitlilik gösterir.
Numara:
Hücre başına Golgi yığınlarının sayısı, hücre tipine bağlı olarak, yüzlerce kişiden yüzeye kadar değişmektedir. Bazı hücrelerde tek bir büyük, Paramoeba durumunda iki tane vardır. Stereomyxa'da (bir Amip türü) birçok Golgi kompleksi vardır. Sinir hücreleri, karaciğer hücreleri ve çoğu bitki hücresi, karaciğer hücrelerinde yaklaşık 50 olan birçok Golgi kompleksine sahiptir.
Algal rizoitlerinde 25.000'den fazla Golgi kompleksi görülür (Sievers, 1965). Golgi kompleksi, bazı özel hücrelerde hücre hacminin büyük bir kısmını bile hesaba katabilir. Bir örnek, bağırsağın içine mukus salgılayan bağırsak epitelinin kadeh hücresidir; mukustaki glikoproteinler esas olarak Golgi kompleksinde glikosile edilir.
Boyut:
Boyut aynı şekilde değişkendir. Sinir ve salgı hücrelerinde büyük, kas hücrelerinde küçüktür. Genel olarak, Golgi kompleksi, hücre aktif durumdayken iyi gelişmiştir. Hücre yaşlandıkça, kompleks aşamalı olarak küçülür ve kaybolur.
Pozisyon:
Golgi kompleksinin pozisyonu, her hücre tipi için nispeten sabittir. Ektodermal kaynaklı hücrelerde Golgi kompleksi, çekirdek ve çevre arasındaki embriyonik durum zamanından polarize olur (Cajal, 1914). Genelde tipik bir polarizasyonu olan salgılayıcı ekzokrin hücrelerinde, Golgi kompleksi, çekirdek ve salgı kutbu arasında bulunur.
Endokrin bezlerinde bu organoid kutupları, folikülün merkezine doğru yönlendirildiği tiroid hariç, değişkendir. Genç hücrelerde ve sıklıkla eski hücrelerde en sık çekirdeğin bir tarafında bulunur, ancak bazı durumlarda onu tamamen sarabilir. Farenin ganglionik hücrelerinde pozisyon perinükleerdir.
Golgi Kompleksi'nin Ayrıntılı Yapısı:
Dalton ve Felix (1954), ilk elektron mikrograflarını aldıktan sonra sıçanın epididiminde Golgi kompleksini tarif etmişlerdir.
Golgi kompleksinin aşağıdaki açıklaması, birkaç yazarın eserine dayanan karma bir tanımdır:
1. Sarnıç :
Sarnıçlar veya kesikler, pürüzsüz yüzey ER'ye benzer ve bölümler, yakından yerleştirilmiş zarla sınırlanmış keseler yığınları halinde görünür. Saccules sayısı çoğu hayvan ve bitki hücresi tipinde 4-8 arasında değişmektedir. Euglena'da, sayı 20'ye kadar çıkabilir.
Saksıların zarı, kenarları çoğunlukla dilate edilen yaklaşık 150 ° C genişliğindeki bir boşluğu çevreleyen yaklaşık 60 ila 70 ° C arasındadır. Çoğu yazara göre, sarnıçların iyi tanımlanmış iki yüzü vardır; yani dışbükey ve içbükey; Sonuncusu genel olarak olgun veya biçimlendirici veya uzak yüz olarak adlandırılır ve dışbükey tarafın olgunlaşmamış veya uyarıcı veya yakın yüz olduğu varsayılır, paralel dizideki sisteran yalanı yaklaşık 200 ila 300 A'lık bir boşlukla birbirinden ayrılır. 0 .
Onları bir arada tutan şey henüz bilinmemektedir, ancak birkaç hücrede, bazı bölgelerde daha belirgin olan kılçıklar arasında Amos ve Grimstone (1968) düğümlerini uygulayan ince kesikli elektron opak tabakası görülür. Mollenhauer ve arkadaşları, (1973), bazı Golgi kompleksi bitkilerinde, intercisternal unsurları ve plakları detaylı bir şekilde araştırdılar.

2. Borular :
Cisterna'nın periferik bölgesinden, 300 ila 500 ° C çapında bir karmaşık, anastomoz yapan düz boru ağı ağı ortaya çıkar. Palyaçolar ve Ardıç (1969) bu boru şeklindeki ağı dantel diskiyle karşılaştırmıştır.
3. Vesiküller :
Veziküller, sarnıç çevresinin çevresinde borulara bağlı kalan küçük damlacık benzeri keselerdir.
Aşağıdaki iki tiptedirler:
(a) Pürüzsüz veziküller :
Düz veziküller 20 ila 80µ çaptadır. Bunlar, salgı malzemesi içerirler (sıklıkla salgı vezikülleri olarak adlandırılırlar) ve ağ içindeki sarnıç boruların uçlarından tomurcuklanırlar. Çoğunlukla birden fazla tübül bağlantısı yapılır ve muhtemelen tek bir oluşturma keseciği doldurun.
(b) Kaplanmış veziküller :
Kaplanmış veziküller, yaklaşık 50 um çapında ve pürüzlü bir yüzeye sahip küresel çıkıntılardır. Organellanın çevresinde, genellikle tek tübüllerin uçlarında bulunurlar ve salgı veziküllerinden morfolojik açıdan oldukça farklıdırlar. İşlevleri bilinmemektedir.
4. Golçya vakuolleri :
Bunlar Golgi'nin olgunlaşan yüzünde bulunan büyük yuvarlak keselerdir. Bunlar, genişletilmiş sarnıçlar veya salgı veziküllerinin füzyonu ile oluşturulur. Vakumlar amorf veya granüler bir madde ile doldurulur.
Golgi kompleksi yapısal ve Biyokimyasal olarak polarizedir:
Golgi kompleksinin iki farklı yüzü vardır: bir cis veya yüz oluşturan ve bir trans veya olgunlaşan yüz. Cis yüzü, kaba ER'nin yumuşak bir geçiş bölümü ile yakından ilişkilidir. Salgılayıcı hücrelerde, trans yüz plazma zarına en yakın olan yüzdür: burada, büyük salgı vezikülleri sadece bir Golgi istifinin trans yüzü ile ilişkili olarak bulunur ve bir salgılama keseciğinin membranı genellikle son ("en çok") cisterna'nın trans yüzü.
Buna karşılık, küçük Golgi veziküllerinin yığın boyunca daha dengeli bir şekilde lokalize olduğu, proteinlerin genellikle cis tarafındaki ER'den bir Golgi yığınına girdiği ve trans tarafındaki çoklu hedefler için çıkış yaptığı düşünülmektedir; bununla birlikte, ne Golgi kompleksi boyunca tam yollarını ne de ne yığınıyla sarnıçtan sarnıçlara gittiklerini bilmiyorlar.
Golgi kompleksinin iki yüzü biyokimyasal olarak belirgindir. Örneğin, Golgi membranlarının kalınlığındaki bir değişiklik bazı durumlarda istif boyunca algılanabilir, cis tarafındakiler daha ince (ER benzeri) ve trans tarafındakiler daha kalın (plazma zarı benzeri).
Daha çarpıcı olan, bazı histokimyasal testlerin Golgi kompleksi içindeki belirli proteinleri lokalize etmek için elektron mikroskobu ile birlikte kullanıldığında elde edilen sonuçlardır. Bu testlerin bazıları membran bağlı enzimi, Golgi yığını içindeki lokalizasyonlarında belirgin bir kutupsallık gösteren aktiviteler ortaya koymaktadır.
Özellikle merak uyandıran bir biyokimyasal bulgu, asit fosfataz gibi lizozomal enzimlerin Golgi istifinin en trans-cisternaları ile ve yakındaki kaplanmış veziküllerin bir kısmı içinde yoğunlaştığını keşfetti. Bu, lizozomlar için ayrılan spesifik veziküllerin bu bölgede toplandığını gösterir.
Sekreter proteinler, istiflenen tüm cisternalarda histokimyasal yöntemlerle bulunur, bu ürünlerin yoğunlaştığı ve sadece trans-en Golgi cisterna ile bağlantılı olduğu büyük salgı vezikülleri.
Kimyasal bileşim:
Golgi kompleksinin kimyasal bileşimi ile ilgili olarak, aşağıdaki maddelerin mevcut olduğu gösterilmiştir:
1. Fosfolipitler :
Golgi membranlarının fosfolipitlerinin bileşimi, endoplazmik membranlarla plazma zarlarının arasındadır.
2. Proteinler ve Enzimler :
Farklı bitki ve hayvan hücrelerinden gelen Golgi kompleksi, protein ve enzim içeriğinde büyük farklılıklar gösterir. Enzimlerin bazıları ADPase, ATPase, NADPH sitokrom-C-redüktaz, glikosi 1 transferazlar, galaktoksi 1 transferaz, tiamin pirofosfat vs.'dir.
3. Karbonhidratlar :
Hem bitki hem de hayvan hücrelerinin glukosarin, galaktoz, glukoz, mannoz ve fruktoz gibi bazı ortak karbonhidrat bileşenleri vardır. Bitki Golgi sialik asit eksikliği, ancak sıçan karaciğerinde yüksek konsantrasyonda ortaya çıkar. Xylulose ve arabinoz gibi bazı karbonhidratlar sadece bitki hücrelerinde bulunur.
4. Vitamin С :
Golgi kompleksinde depolanan С vitamini oranı Tomitte tarafından gösterilmiştir. Ona göre Golgi kompleksi С vitamini depolar ve hücre ürünlerinin oksidasyonunu önlemek için yavaşça sitoplazmaya serbest bırakır.

Golgi kompleksinin fonksiyonları:
1. Spermiyojenez sırasında Akrozomun Oluşumu :
Sperm olgunlaşması sırasında Golgi kompleksi akrozom oluşumunda rol oynar (Burgos ve Fawcett, 1955).

Golgi'nin erken evrelerinde, paralel istifler ve çok sayıda küçük veziküller şeklinde düzenlenmiş sarnıçlardan oluşan küresel bir gövde olarak görülür. Daha sonra hep sarnıçtan ayrıldı. Gelişme ilerledikçe, Golgi kompleksi düzensiz hale gelir ve cisternal keselerin açılımıyla büyük boşluklar oluşur.
Bu büyük vakuol veya vakuollerin merkezinde proakrozomal granül olan yoğun bir granül bulunur. Golgi kompleksinden türetilen bu granül, vakum olarak bilinen bir işlemle vakum içinde büyümeye devam ediyor. Bu vakuol ve granül, akrozomal granül oluşturan nükleer membranın ön kutbuna yaklaşır.
Spermatidin uzamasıyla, akrozomal vezikül nükleer yüzeye yayılır ve nükleer zarla çökerek başlık materyalini oluşturur. Akrozomal granül, çekirdeğin tepesinde bulunan akrozom haline gelir ve görünüşe göre döllenme işleminde yer alan bazı enzimleri içerir.
2. Polisakaritlerin sentezi ve salgılanması:
Kadeh hücrelerinde otoradyografi ve elektron mikroskobu ile yapılan çalışmalar protein sentezi, karbonhidrat ilavesi ve sülfaj arasındaki ilişkiyi kurmuştur. Kolonun goblet hücreleri mucigen üretir. Bu salgılama maddesi büyük oranda karbonhidrat içerir.
Golgi kompleksi çekirdeğin hemen üstünde bulunur. Hücrenin serbest yüzeyine doğru yavaş yavaş mucigen granülleri büyüyor. Golgi kompleksinin proksimal sarnıçları herhangi bir şişlik göstermez, ancak yığın boyunca belirli bir mesafede distal sarnıçları aniden mucigen granüllerine dönüştürülür. Distal cisternae sürekli olarak her 2-4 dakikada bir mucigen granüllerine dönüşür. Yeni proksimal cisterna tazminat oluşur.
3. Salgılamadaki rol:
Golgi kompleksinin bir hücrenin salgılama fonksiyonunda bir rol oynadığı düşünülmektedir. Ancak asıl soru, bazı maddeleri kendileri salgıladıkları veya sentezledikleri ya da hücrede başka bir yerde salgılanan salgı ürünlerinin basit bir şekilde depolandığı ve konsantre oldukları bir depo olduklarıdır.


Palade ve ark. 1962 bu salgı döngüsü, şimdi iyi tanımlanmıştır ve pankreas asiner hücrelerinde dört adım içermektedir ve bunlar:
(i) Amino asitlerin kaba endoplazmik retikulum yüzeyindeki proteine dahil edilmesi.
(ii) Bu yeni salgılayıcı proteinlerin kaba endoplazmik retikulum sarnıçlarına aktarılması.
(iii) Bu proteinlerin hücre içi Golgi kompleksine taşınması.
(iv) Zimogen granüllerin, lümenlere boşaldıkları hücrenin tepesine doğru göçü.
4. Gologi gövdesinin oogenezdeki rolü :
Srivastava (1965), Gologen kompleksi hakkında, oogenesis sırasında kısa bir inceleme yapmıştır. Afzelius'a (1956) göre, elektron mikroskobu altında görüldüğü gibi deniz kestanesi yumurtasının Golgi kompleksi, zaman zaman şişebilen düz torbaların lamel oluşturan duvar yığınlarından oluşur.
Bu organların enine bölünmelerinin bazı belirtileri vardır. Sotelo (1959) ve Sotelo ve Porter (1959), elektron mikroskobu altında görüldüğü gibi sıçan-ovumdaki Golgi kompleksini tarif etmiş ve bu organellerin erken oositlerde juxtra nükleer yerleşimini bulmuşlardır.
Bir sonraki aşamada, bunlar parçalara ayrılır ve üçüncü aşamada, bunlar kortekse doğru hareket eder. Tüm bu durumlarda, yapıları, ince, çift profilli (düzleştirilmiş keseler) ve küresel veziküllerin yakın paketlenmiş dizileri olarak kalır.
Erken oositlerde, kompleks kompakt bir şekilde düzenlenmiştir. Daha sonraki aşamalarda, kortikal bölgeye dağılmış küçük veziküller ile çevrili ayrı profil demetleri bulunur. Erken oositlerde, Golgi kompleksi ve centrozomu yakından ilişkilidir.
5. Bileşiklerin absorpsiyonu :
Hirsch ve ark., Demir şekeri bir hayvana beslendiğinde demirin Golgi kompleksinde (Kedrowsky) emildiğini keşfetti. Van Teel, Golgi sistemlerinin aynı zamanda bakır ve altın bileşiklerini de emdiğini göstermiştir. Kedrowsky, Opalina'nın Golgi kompleksinin bismutoz (albümin ve bizmut bileşiği) ve protargol (albümin ve gümüş bileşiği) absorbe edebileceğini göstermiştir. Böylece, Kirkman ve Severinghaus, Golgi kompleksinin ürünlerin damlacıklara veya granüllere konsantrasyonu için yoğunlaşma zarı olarak davrandığını belirtmektedir.
6. Bitki hücre duvarı oluşumu :
Bitkilerin hücre duvarı, bazı lipidler ve proteinlerle birlikte, ağırlıklı olarak polisakaritler içeren fibrillerden oluşur. Sitokinez sırasında, iki kız çekirdeği arasında bir hücre plakası oluşturulur ve çevresinde daha sonra kız hücrelerinin plazma zarı haline gelen bir zara sahiptir. Polisakkaritlerin Golgi kompleksinde oluştuğuna ve hücrelerin hala büyürken ortaya çıkan yeni hücre duvarına aktarıldığına dair net kanıtlar vardır.

Plazma zarlarını ayıran hücre plakasının matrisini oluşturan pektinler ve hemiselülozlar gibi maddeler de Golgi kompleksinden katkıda bulunur.
7. Hücre içi kristallerinin oluşumu :
Deniz izopodunda, oyuk haline getirilmiş bir form olan Limnoria lingmorum, hücreleri kristallerden oluşan orta yaylalarda bulunur. Bunlar, 30 ° C uzunluğa ve 15 ° C kalınlığa kadardır. Bu kristallerin Golgi kompleksi tarafından oluşturulduğu ve protein ve demir içerdiği bilinmektedir. Onlar zarfı kapatamazlar ve genellikle sferoidal formdadırlar. Salgı aktivitesiyle ilgileniyorlar.
8. Süt Protein damlacık oluşumu :
Emziren memenin farelerinde, Golgi kompleksi ile ilgili protein damlacıkları üretilir. Bu damlacıklar, genellikle hücre zarlarına, kendi zarlarının plazma membranı ile birleştirilmesiyle açılırlar.
9. Lizozom ve vakuollerin oluşumu :
Birincil lizozomlar, Golgi membranlarından salgılayıcı veziküller ile aynı şekilde oluşturulur. Diktiyomların, daha olgun bölgelerinde hidrolitik enzimler biriktirdiğine dair iyi kanıtlar vardır. Bitki hücrelerinde bulunan bazı vakuollerin az miktarda hidrolitik enzim içerdiği ve bunların Golgi kompleksinden geldiği tahmin edilmektedir.
10. Pigment oluşumu:
Birçok memeli tümör ve kanser hücresinde Golgi kompleksi, pigment granüllerinin (melanin) orijini bölgesi olarak tarif edilmiştir.
11. Akışkan dengesinin düzenlenmesi :
Golgi kompleksi ve alt Metazoa ve Protozoa'nın kasılma vakuolü arasında bir homoloji öne sürülmüştür. Kasılma vakuolü fazla suyu hücreden atıyor. Bazı Protozoa'larda Golgi kompleksi ayrıca sıvı dengesinin düzenlenmesiyle de ilgilidir.
Golgi Kompleksinin Kökeni:
Yeni Golgi kompleksinin ortaya çıkabileceği üç farklı kaynak önerildi:
1. Endoplazmik retikulumdan :
Essner ve Novik®ff (1962) ve Beams and Kessel (1968) Golgi cisternae'nin ER'den kaynaklandığını öne sürdüler. Spesifik proteinleri sentezledikten sonra kaba endoplazmik retikulum ribozomları kaybeder ve yumuşak ER'ye dönüşür. Küçük geçici veziküller pürüzsüz ER'den ayrılırlar. Bunlar diktiyomere göç eder.
Bu sigorta dityozomun şekillendirici yüzüne ulaştığında yeni sarnıçlar oluşturur ve böylece büyümesine katkıda bulunur. Bu veziküllerin füzyonu ile sürekli olarak yeni cisterna şekillendirme yüzünde oluşur ve olgunlaşan yüzünde eski cisterna salgı veziküllerine ayrılır. Böylece Golgi membranöz akış fenomeni sergiler. '
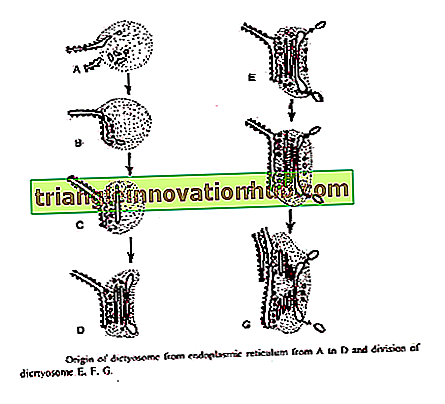

Bouch (1965), kahverengi alglerdeki nükleer zarfın dış zarından Golgi'nin kökenini tanımlamıştır. Veziküller diktyozomun şekillendirici yüzünde cisterna oluşturmak için kaynayan dış nükleer zardan sıkıştırılır.
Pürüzsüz ER veya nükleer membran ile ilişkili olarak dışlama bölgelerinin varlığı, daha yüksek bitkilerin hareketsiz tohumlarında dışlama bölgelerinin oluşması ve çimlenen tohumlarda bu bölgelerden diktiyom oluşumu, tohumların kökeni hakkındaki iki teoriyi destekleyen kanıtlar sunar. dictyosome.
3. Önceden var olan diktiyom bölünmesi ile :
Hem bitkilerde hem de hayvanlarda hücre bölünmesi sırasında, dityozomların sayısının arttığı ve bölünmeden hemen sonra her bir kız hücresindeki diktiyomların sayısının, bölünmeden hemen önce ana hücredeki sayıya, teneke ve diğer direklere eşit olduğu gözlenmiştir. bölünen hücreler üzerinde yapılan gözlemler, diktiyomların hücre bölünmesi sırasında da bölündüğü varsayılmıştır.